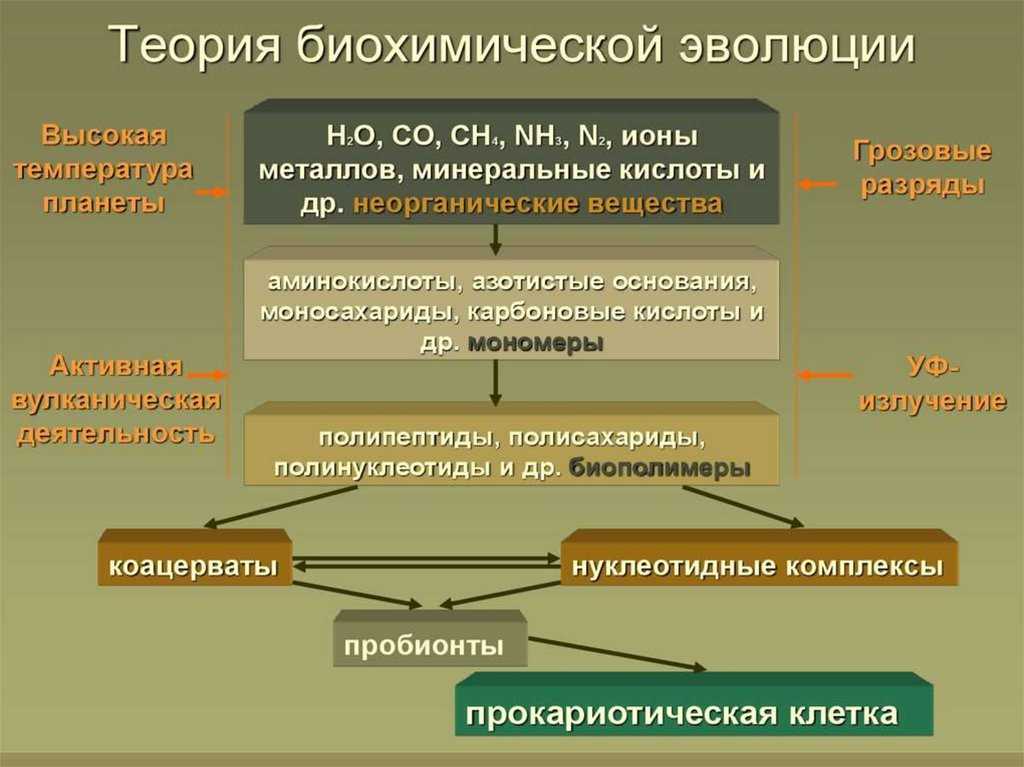
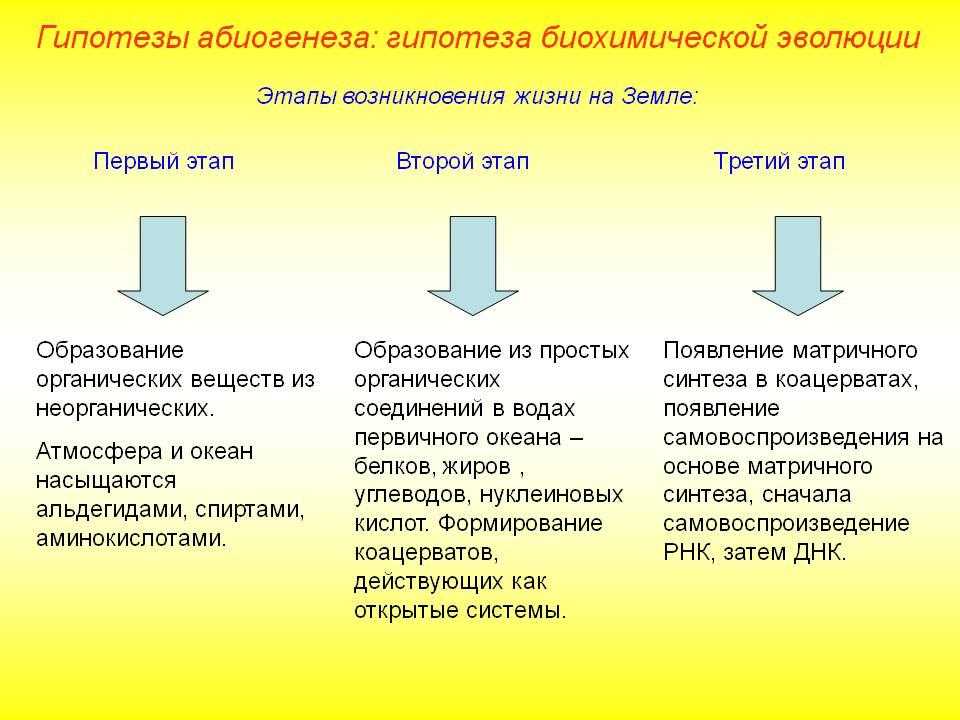
ВВЕДЕНИЕ
Система антиоксидантной (АО) защиты – один из важнейших элементов формирования адаптаций
гидробионтов в условиях окислительного стресса (ОС). Эта система обеспечивает неспецифический
отклик организма на окислительный стресс, возникающий при действии самых разных экологических
и антропогенных факторов в водной среде, ее показатели широко применяются в оценке
состояния здоровья водных организмов и среды в целом . Двустворчатые моллюски-фильтраторы – одни из наиболее устойчивых гидробионтов к
действию ОС разного генеза. Выявлена высокая способность двустворчатых моллюсков к
детоксикации активных форм кислорода (АФК) , что связывают с эффективностью их защитных систем, и в том числе АО системы. Состояние
АО комплекса и перекисного окисления липидов (ПОЛ) двустворок зависит как от эндогенных
процессов, так и от факторов внешней среды . Особенности системы АО защиты и соотношение ее показателей с уровнем ПОЛ в значительной
мере определяют устойчивость этих животных к окислительному стрессу . В этой связи актуально сравнительное исследование АО комплекса и ПОЛ у двустворчатых
моллюсков с высокой устойчивостью к неблагоприятным факторам, – в частности, у черноморских
мидии Mytilus galloprovincialis, анадары Anadara kagoshimensis, сердцевидки Cerastoderma glaucum. Это массовые эврибионтные моллюски, широко распространенные в Азово-Черноморском
бассейне и других районах Мирового океана, играющие важную роль в морских экосистемах
.
Моллюски рода Cerasroderma отличаются высокой выживаемостью в неблагоприятных условиях, выдерживают значительные
колебания солености – 5–38‰ и температуры – 0–25°С, а также гипоксию, обсыхание и
другие неблагоприятные воздействия . C. glaucum населяет биотопы с высокой загрязненностью [], превосходит других бентосных животных по устойчивости к загрязнению тяжелыми металлами
, органическими поллютантами [], что во многом связывают с высокой активностью АО ферментов СОД и каталазы .
Анадара длительно выживает при аноксии , в среде с крайне низким уровнем кислорода в течение 5–7 дней []. АО комплекс анадары более эффективен по сравнению с мидией и устрицей Черного моря,
что выражается в более высоком содержании глутатиона и активности глутатионпероксидазы
в ноге, повышенной активности СОД и каталазы в жабрах и в более низком уровне ПОЛ
во всех исследованных тканях , а также активном участии в АО защите анадары как ферментного, так и низкомолекулярного
звена [].
Мидия Mytilus sp. достаточно устойчива к действию широкого ряда поллютантов, комплексного влияния загрязнения
и гипоксии, включена в программы биомониторинга для оценки качества водной среды,
для чего разработан ряд широко применяемых биомаркеров, таких как СОД, каталаза, глутатионтрансфераза
. В гипоксических условиях мидия способна переходить к анаэробному обмену, хотя и
при более высоком уровне кислорода в среде, чем для анадары [].
Состояние АО комплекса и ПОЛ у моллюсков в естественных условиях обитания имеет, как
правило, выраженную тканевую и видовую специфику . Так, установлены специфические особенности АО системы и ПОЛ у мидии, анадары и
устрицы Черного моря , ряда видов дальневосточных моллюсков [], анадары и мидии Средиземного моря []. Вызывает интерес и то, что выраженные различия АО параметров показаны и у моллюсков
одного и того же или близких видов – в тканях трех цветовых морф мидии M. galloprovincialis [], двух видов дрейссен из Рыбинского водохранилища []. У близкородственных видов сердцевидок C. glaucum и C. edule были выявлены различия в потреблении ими кислорода, а также в поведении и выживаемости
в экстремальных условиях при обсыхании, аноксии []. В отношении влияния загрязнения подчеркивается, что разная аккумулирующая способность
(на примере трех видов моллюсков C. edule, Crassostrea gigas и Ruditapes philippinarum) также может быть связана с физиологическими различиями между этими видами и может
служить индикатором стресса у этих животных [].
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
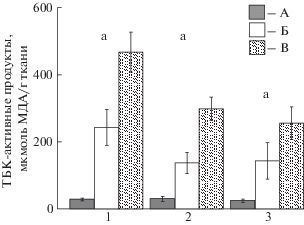
Сердцевидка отличалась наименьшим содержанием ТБК-активных продуктов – во всех исследованных
тканях эта величина была в 4.5–16 раз ниже (p ≤ 0.001), чем у анадары и мидии (). Значение этого показателя уменьшалось в ряду моллюсков мидия – анадара – сердцевидка.
Наибольшие отличия наблюдались между сердцевидкой и мидией – в 10–16 раз (p ≤ 0.001), при этом наиболее выраженные – в гепатопанкреасе, а наименьшие – между
мидией и анадарой – 1.8–2.1 раза (p ≤ 0.05–0.01) (все ткани).
Рис. 1.
Содержание ТБК-активных продуктов в тканях сердцевидки Cerastoderma glaucum, анадары Anadara kagoshimensis, мидии Mytilus galloprovincialis (1 – гепатопанкреас, 2 – жабры, 3 – нога; А – сердцевидка, Б – анадара, В – мидия;
отличия достоверны при p ≤ 0.05–0.001: a – между всеми видами, b – между сердцевидкой и анадарой, c – между
сердцевидкой и мидией, d – между мидией и анадарой).
Наибольшую активность обоих исследованных ферментов зарегистрировали в жабрах сердцевидки.
Активность СОД была в 5.8–11.6 раза выше (p ≤ 0.001) по сравнению с жабрами двух других видов (). При этом, как и в случае ТБК-активных продуктов, наиболее выраженные отличия найдены
между сердцевидкой и мидией (11.6 раза) (p ≤ 0.001), в то время как жабры мидии и анадары отличались по этому показателю только
вдвое (p ≤ 0.05).








Рис. 2.
Активность СОД в тканях сердцевидки Cerastoderma glaucum, анадары Anadara kagoshimensis, мидии Mytilus galloprovincialis. Обозначения – как на рис. 1.
В гепатопанкреасе сердцевидки активность СОД превышала таковую у анадары в 3.3 раза
(p ≤ 0.05), а у мидии – в 1.5 раза, но в этом случае отличия не были достоверными. Сердцевидка
отличалась наиболее высокой активностью СОД и в ноге – в 2.1 и 4.7 раза больше (p ≤ 0.05–0.01), чем у мидии и анадары соответственно.
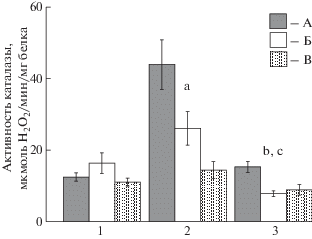
Самая высокая активность каталазы из трех исследованных тканей зафиксирована в жабрах
моллюсков каждого из видов, она снижалась в ряду сердцевидка – анадара – мидия (). Как и в случае других показателей, наибольшие отличия по каталазе в жабрах обнаружены
между сердцевидкой и мидией – в 3.1 раза (p ≤ 0.01), а между остальными моллюсками – в 1.7–1.8 раза (p ≤ 0.05). Активность фермента была выше у сердцевидки и в ткани ноги – в 1.7–2.0 раза
(p ≤ 0.05) по сравнению с особями других двух видов. В отличие от жабр и ноги, в гепатопанкреасе
моллюсков всех исследованных видов каталаза проявляла близкую активность, значимых
отличий не установлено.
Рис. 3.
Активность каталазы в тканях сердцевидки Cerastoderma glaucum, анадары Anadara kagoshimensis, мидии Mytilus galloprovincialis. Обозначения – как на рис. 1.
Для дополнительной оценки эффективности системы СОД-каталаза было рассчитано соотношение
величин активности этих ферментов (). Наиболее высокие значения коэффициента отмечены у сердцевидки во всех исследованных
тканях. В жабрах сердцевидки коэффициент в 3.5–3.8 раза выше, чем у анадары и мидии,
а в гепатопанкреасе – в 4.4 раза больше по сравнению с анадарой. В остальных случаях
отличия были меньше – в 1.4–2.4 раза.
МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЙ
Материалом исследования служили половозрелые особи двустворчатых моллюсков – сердцевидки
C. glaucum (Bivalvia: Cardiidae), анадары A. kagoshimensis (Bivalvia: Arcidae) и мидии M. galloprovincialis (Bivalvia: Mytilidae) с длиной раковины 26–29, 30–34 и 55–58 мм соответственно. Моллюски
были собраны в начале–середине мая 2018 г. в прибрежной зоне бухты Казачья (район
Севастополя, Черное море). Особей каждого вида собирали в их природных местообитаниях:
сердцевидку – на мелководье, в нескольких метрах от уреза воды, на глубине 0.5–0.7
м на илисто-песчаном грунте, а анадару и мидию – в более глубокой части бухты на глубине
2.0–2.5 м; анадару собирали со дна, а мидию – как со дна, так и с твердых субстратов
(прибрежные скалы). Особей сердцевидки и анадары выкапывали с помощью перфорированной
емкости, так как моллюски были немного погружены в грунт. Температура воды на малой
глубине, где собирали сердцевидку, составила 17.4°С, а на большей глубине, где обитали
анадара и мидия, – 16.6°С. Концентрация кислорода для этих местообитаний составила
0.85–0.9 мл О2/л и 2.9–3.1 мл О2/л соответственно. Измерения проводили с помощью кислородомера Starter 300D (производство
Ohaus, США).








Известно, что показатели АО комплекса и ПОЛ весьма чувствительны к сезонным колебаниям
параметров водной среды – температуры, концентрации кислорода, солености, содержания
поллютантов и др., а также к изменениям в физиологическом состоянии моллюсков и, в
первую очередь, к состоянию нереста . Как было показано нами ранее [], физиолого-биохимические изменения в организме мидии при нересте оказывают влияние
на состояние АО комплекса и ПОЛ моллюска – в большей степени гепатопанкреаса, чем
жабр. Многие двустворчатые моллюски относятся к видам, нерестящимся круглогодично
с большей или меньшей интенсивностью. Однако наиболее активный нерест у моллюсков
происходит в определенные сезоны года – пики нереста: у мидии – в марте–апреле и октябре–ноябре
[], у анадары – с июня по сентябрь [], у сердцевидки интенсивный гаметогенез – в марте–начале апреля, а первый вымет гамет
– в середине мая, второй – в августе []. Поэтому для унификации величин абиотических факторов, а также во избежание влияния
активного нерестового состояния на АО комплекс и уровень ПОЛ в тканях моллюсков, их
собирали почти одновременно в короткий промежуток времени в начале–середине мая: особей
сердцевидки – в самом начале мая, до начала активного вымета гамет; мидию и анадару
– в середине мая, когда у этих моллюсков еще не наступает пик нереста.
После сбора моллюсков содержали в аквариумах с проточной морской водой в течение двух
суток для снятия стресса после транспортировки. Исследуемые ткани моллюсков – гепатопанкреас,
жабры и мышечную ткань (ногу) – гомогенизировали при температуре 0–4°С с использованием
ледяной бани. Гомогенаты центрифугировали при 3200 g в течение 15 мин с помощью рефрижераторной
центрифуги К-23D (Германия). В приготовленных супернатантах определяли активность
АО ферментов – супероксиддисмутазы (СОД) по реакции ингибирования восстановления нитросинего
тетразолия в присутствии НАДН и феназинметасульфата, длина волны 540 нм []. Активность каталазы измеряли по интенсивности желтой окраски в результате реакции
остаточных количеств пероксида водорода (после взаимодействия с ферментом) с молибдатом
аммония при длине волны 410 нм []. Активность СОД выражали в мкмоль НАДН в минуту на 1 мг белка, каталазы – в мкмоль
пероксида водорода в минуту на 1 мг белка. Интенсивность перекисного окисления липидов
(ПОЛ) оценивали по накоплению продуктов, реагирующих с тиобарбитуровой кислотой (ТБК-активных
продуктов), при длине волны 532 нм, их содержание выражали в микромолях малонового
диальдегида (МДА) на 1 г сырой ткани []. Содержание белка определяли по методу Лоури с реактивом Фолина–Чокальтеу [].
Таксономия
Таксон, вероятно, впервые был описан Ловеллом Августом Ривом в 1844 году как Arca inflata . Однако этим названием занимались дважды: Arca inflata Schroeter, 1802 и Arca inflata Brocchi, 1814. В 1848 году Нист предложил заменить название Arca reeveana , которое было дано Arca reeveana d’Orbigny, 1844. Его заменяет название Arca broughtonii, созданное Леопольдом Шренком в 1867 году . В настоящее время этот вид является частью рода Anadara Grey, 1847, в качестве альтернативы некоторыми авторами подроду Anadara (Scapharca) Gray, 1847. Подрод рода Anadara не является общепринятым.
Старые синонимы — Arca inflata Reeve, 1844 (недействительны, потому что младший омоним Arca inflata Brocchi, 1814) и Arca reeveana Nyst, 1848 ( альтернативное название для Arca inflata Reeve, 1844, но недействителен, потому что младший омоним Arca reeveana d’Orbigny, 1846) ). Более поздние синонимы: Arca tenuis Tokunaga, 1906 (не Arca tenuis Montagu, 1808) и Anadara kafanovi Lutaenko, 1993.
функции
Корпус с одинаковыми петлями имеет длину 13,6 см, высоту 11,4 см и толщину 10,8 см (до 15 см в длину). Он явно неравный, позвонки находятся в передней половине корпуса. Доральный край прямой, образует с передним краем угол от почти прямоугольного до слегка тупого. Передний край выглядит немного обрезанным до очень плоского. Угол дорсального края с задним краем пологий. Затем задний край ниспадает прямо вниз до плотно закругленного перехода к брюшному краю. Вентральный край хорошо закругленный. Позвонки широкие, загнутые вперед. Они относительно далеки друг от друга.
Пластина замка имеет прямой верхний край и умеренно вогнутый нижний край. Поэтому стопорная пластина довольно узкая посередине. Таксодонтический замок состоит из множества вертикальных зубцов, которые сначала становятся выше кнаружи и немного расходятся кнаружи. Крайние зубы часто имеют угловатую форму.
Раковина сравнительно тонкая для своего размера. Он грязно-белый, от светло-коричнево-желтого до коричневатого оттенка, иногда также темно-коричневый. Внутри скорлупа беловатого цвета со светлым желто-коричневым оттенком; чистый белый цвет между линией поверхности и краем корпуса. Имеется от 42 до 43 радиальных уплощенных ребер. Ребра немного шире, чем промежутки и углубления между ребрами. Они пересекаются линиями роста, которые вызывают небольшие, лишь слегка глубокие выемки в точках пересечения с ребрами или, реже, несколько зернистые в точках пересечения. В ювенильных частях корпуса линии роста видны только в промежутках между ребрами, в краевых областях также на ребрах. Периостракум темно — коричневого до черновато — коричневого и отслаивается легко. В основном он сохранился только на краю корпуса и в углублениях между ребрами жесткости. Он состоит из длинных потертых, часто скатывающихся чешуек.
Ребра и впадины снаружи также отмечены изнутри слабыми радиальными выступами и впадинами. На внутренней стороне также видны очень тонкие радиальные полосы частично приподнятых и частично углубленных линий, за исключением точек прикрепления мышц сфинктера. Полосы оканчиваются на поверхности. Есть два сфинктера почти одинакового размера.