

Примечания
Комментарии
-
Филогенетические определения:
- Eotetrapodiformes: ближайший общий предок Eusthenopteron и Ichthyostega и все его потомки;
- Стегоцефалы (Stegocephali): наибольшая клада, включающая Eryops megacephalus, но не Tiktaalik roseae, Panderichthys rhombolepis и Eusthenopteron foordi;
Источники
- ↑ , с. 625.
- Ископаемые позвоночные России и сопредельных стран. Бесчелюстные и древние рыбы. Справочник для палеонтологов, биологов и геологов / Под ред. Л. И. Новицкой, гл. ред. акад. Л. П. Татаринов. — М.: ГЕОС, 2004. — С. 271. — 436 с. — ISBN 5-89118-368-7.
- ↑ , с. 190.
- Палеонтология и совершенствование стратиграфической основы геологического картографирования. — Санкт-Петербург: Материалы LV сессии Палеонтологического общества при РАН (6—10 апреля 2009 г. Санкт-Петербург), 2009. — С. 110. — 184 с.
- , с. 152, 625.
- , с. 161, 163—164.
- , с. 163, 190.
- ↑ , с. 143.
- Кэрролл Р. Палеонтология и эволюция позвоночных: В 3-х тт. Т. 3. — М.: Мир, 1993. — 312 с. — ISBN 5-03-001819-0. — С. 190—191.
- , с. 625—634.
- , p. xi—xii, 102—111.
- , p. 103.
- Merck J. (англ.). GEOL 431 Vertebrate Paleobiology. Дата обращения: 1 июля 2021.
- — S2CID .
- , Stegocephali E. D. Cope 1868 , converted clade name, pp. 741—745.
- (англ.). RegNum. Дата обращения: 29 июня 2021.
- , p. 629.
- Марков А., Неймарк Е. Эволюция. Классические идеи в свете новых открытий. — М.: ACT: Korpus, 2014. — С. 486. — 656 с. — ISBN 978-5-17-083218-7.
- , p. 634.
- ↑ , p. 626.
Надотряд кистепёрых рыб
У одной группы кистеперых, относимой к отряду рипидистиеобразных (Rhipidistiiformes),
эволюция пошла в направлении приспособления к дыханию атмосферным воздухом (внутренние ноздри, легкие)
и преобразования парных плавников в пятипалые конечности.
Дав начало первым наземным позвоночным и просуществовав около 100 миллионов лет,
рипидистиеобразные кистеперые вымерли.
Другая группа кистеперых, образовавшая отряд целакантообразных (Coelacanthlf ormes),
сначала развивалась преимущественно в пресных водах, затем приспособилась к обитанию в море.
От двух отрядов и 9 семейств кистеперых рыб сохранился один вид — латимерия,
представитель семейства латимериевых отряда целакантообразных.
[Раньше считалось, что именно целакантообразные могли быть предками тетрапод]
Происхождение лопастепёрых рыб
(C) И.К.Гаршин, 2000
Выскажу крамольную гипотезу: и костные рыбы и земноводные произошли от кистеперых.
Сами лопастеперые произошли от хрящевых благодаря переходу к жизни в пересыхающем мелководье.
Сама костная ткань и плавательный пузырь (изначально выполнявшего функцию легкого)
явились приспособлениями к полуназемной жизни.
Затем часть лопастеперых ушла обратно в глубины морские и стала костными рыбами,
часть стала дальше приспосабливаться к суше и стала амфибиями,
другая часть некоторое время продолжала жить между стихиями,
пока ее не вытеснили, наверное, сами амфибии.
Образ жизни протоптеров
Двоякодышащие рыбы проживают в пересыхающих пресных водоемах. В реках они встречаются редко, поскольку предпочитают стоячую воду. В сезон дождей места их обитания затапливаются крупными реками. Протоптеры постоянно подымаются на поверхность, чтобы заглотнуть воздуха. Ученые подсчитали, что животные получают 2 % необходимого кислорода через жабры. Легкие обеспечивают их воздухом на 98 %. А вот молодняку жабры предоставляют до 90 % кислорода.
Протоптеры – ночные охотники. В темное время они намного чаще подымаются для вдоха. Рыбы не только дышат двумя способами, но и передвигаются в воде. Так они могут плавать за счет изгибания тела. Плавники при этом плотно прижаты. Для перемещения по дну они используют плавники.
Рыбы живут в мутной воде, охотятся ночью, поэтому зрение не играет особой роли. Помогают ориентироваться вкусовые почки, которыми усеяны плавники. Важную роль играет обоняние. Днем рыбы довольно вялые и апатичные, они чаще находятся на дне.
В рацион протоптеров входят:
- моллюски;
- пресноводные крабы;
- рыба.
Молодняк до тридцати пяти сантиметров питается насекомыми. Когда взрослая особь нащупывает жертву, она молниеносно нападает на нее, заглатывая ртом. Затем протоптер несколько раз пережевывает добычу. Крупные представители двоякодышащих способны съесть форель. В экстремальных ситуациях они могут долгое время оставаться без пищи. Речь идет о нескольких годах.
Образ жизни лепидосирен
В активное время суток они охотятся, перемещаясь по дну. Их любимой пищей являются моллюски ампулярии. Рацион также дополняют мелкие рыбки и растительность.
Половой зрелости двоякодышащая рыба достигает в три года. Начало нереста припадает на третью неделю после возобновления сезона дождей. Нору готовит самец. Ее глубина до полутора метра, а ширина около двадцати сантиметров. Дно покрывается растительностью.
Самка выметывает икринки диаметром до семи миллиметров. Через две недели из них появляются личинки. Потомство охраняет самец. Через тридцать-сорок пять дней у личинок рассасывается желточный мешочек. Когда они покинут гнездо, их наружные жабры исчезнут. Кормиться первое время они могут водорослями, планктоном.
Протоптеры
Все четыре вида этих двоякодышащих рыб обитают в водах Африки. Внешне они очень похожи, отличаются лишь окрасом и количеством ребер. Обладают вытянутой формой с и жгутообразными плавниками. Особенность этих представителей – при пересыхании водоема впадать в спячку.
Образ жизни
В реках встречается крайне редко, протоптеры предпочитают пресную и мутную воду. Ведут ночной образ жизни, днем вялые, можно найти на дне.
Могут двигаться двумя способами: используя свои плавники для передвижения по дну и плавая, извиваясь телом, прижимая плавники. Для ориентации в пространстве у протоптеров имеются рецепторы в плавниках и развито обоняние, зрение развито слабо.
Эти рыбы питаются ракообразными, пресноводными крабами и рыбой. Молодые рыбки едят насекомых.
Размножение
Двоякодышащие достигают половой зрелости примерно в 3–4 года. Нерест начинается в августе-сентябре, и продолжается в течение одного месяца. В это время протоптеры выходят из спячки.
Взрослая пара строит гнездо с двумя выходами. Пока неясно, кто строит гнездо, так как следить за этими рыбами крайне сложно в естественной среде, а в неволе они не размножаются. Известно, что самцы занимаются охраной яйцекладки, а также содержат мальков. Самцы очень защищают своих детенышей. Через неделю после откладки самкой икры появляются личинки. Они выделяют особый липкий секрет, который помогает им прилипать к стенкам гнезда. Остаются в этом положении до тех пор, пока желточный мешок не рассосется. В это время дыхание происходит с помощью 4 пар наружных жабр.

Через две недели покидают гнездо, подросшие до 2 см, но держатся достаточно близко, чтобы спрятаться в случае опасности. Потомство начинают дышать через легкие и находят себе пищу. Молодь в конце концов покидает гнездо, когда вырастает до 3 см, наружные жабры постепенно исчезают.
Спячка
Уникальное строение протоптера позволяет ему впадать в спячку. Это явление наблюдается только с этим видом. Подготовка к спячке начинается в засушливый сезон.
Если водоем не пересох, рыбы не впадают в спячку. В этом состоянии двоякодышащие проводят до полугода, а в некоторых случаях задерживают до года.
Процесс перехода в спячку начинается с того, что рыба прогрызает ртом дно чтобы добраться до толстого слоя глины и песка. После она заглатывает ил и выбрасывает его жабрами.
Размер укрытия сопоставим с размером рыбы. Внизу делается расширение, где и проводит спячки. В нем рыба складывается пополам, чтобы иметь возможность высунуть голову. Какое-то время рыба активна, пока не преграждает путь глиняной пробкой своим телом, когда поднимается для заглатывания воздуха.

В это время протоптеры выделяют слизь, которая смешивается с водой, становится вязкой и образует кокон. По мере снижения уровня воды в укрытии рыба опускается в укрытие и впадает в спячку. В коконе из слизи и неорганического вещества остается только одно воронкообразное отверстие, соединяющее особь с внешним миром.
Пробуждение в естественных условиях все еще не изучено. В лабораторных условиях рыбы начинают подниматься на поверхность, чтобы заглатывать воздух. Нужно много еды, чтобы восстановится в течение длительного времени.
легкие
Боковой вид легких рассеченной пятнистой двоякодышащей рыбы ( Protopterus dolloi )
Двоякодышащие рыбы имеют узкоспециализированную дыхательную систему . Они имеют отличительную особенность в том, что их легкие соединены с гортанью и глоткой без трахеи. В то время как другие виды рыб могут дышать воздухом с помощью модифицированных васкуляризированных газовых пузырей , эти пузыри обычно представляют собой простые мешочки, лишенные сложной внутренней структуры. Напротив, легкие двоякодышащих рыб подразделяются на множество более мелких воздушных мешочков, что максимально увеличивает площадь поверхности, доступную для газообмена.
Большинство существующих видов двоякодышащих рыб имеют два легких, за исключением австралийских двоякодышащих рыб, у которых только одно. Легкие двоякодышащих рыб гомологичны легким четвероногих . Как и у четвероногих и двуногих , легкие отходят от вентральной поверхности пищевода и кишечника.
Перфузия воды
Из существующих двоякодышащих рыб только австралийские двоякодышащие могут дышать через жабры, не нуждаясь в воздухе из легких. У других видов жабры слишком атрофированы, чтобы обеспечить адекватный газообмен . Когда двоякодышащая рыба получает кислород из жабр, ее кровеносная система устроена так же, как и у обычной рыбы. Спиральный клапан артериального конуса открыт, обходные артериолы третьей и четвертой жаберных дуг (фактически не имеющих жабр) закрыты, артериолы второй, пятой и шестой жаберных дуг открыты, артериальный протокотходящие от шестой артериолы открыты, а легочные артерии закрыты. Когда вода проходит через жабры, двоякодышащие рыбы используют буккальный насос. Поток через рот и жабры однонаправленный. Кровоток через вторичные ламеллы противоточен воде, поддерживая более постоянный градиент концентрации.
Перфузия воздуха
При дыхании воздухом спиральный клапан артериального конуса закрывается (минимизируя смешивание оксигенированной и дезоксигенированной крови), открываются третья и четвертая жаберные дуги, закрываются вторая и пятая жаберные дуги (минимизируя возможную потерю кислорода, полученного в легких). через жабры), артериальный проток шестой артериолы закрывается, а легочные артерии открываются
Важно отметить, что при дыхании воздухом шестая жабра все еще используется при дыхании; деоксигенированная кровь теряет часть углекислого газа, проходя через жабры, прежде чем попасть в легкие. Это связано с тем, что углекислый газ лучше растворяется в воде
Поток воздуха через рот является приливным, а через легкие двунаправленным и наблюдается «равномерная бассейновая» диффузия кислорода.
Слайды и текст этой презентации

Класс Osteichthyes — Костные рыбы
Подкласс Sarcopterygii —Лопастеперые рыбы
Выполнила – Новгородцева А.С.Группа – МЕНМ-170606











тип Chordata — Хордовые подтип Vertebrata (Craniota) — Позвоночные (Черепные)раздел Gnathostomata — Челюстноротыегруппа Anamnia — Первичноводныенадкласс Pisces — Рыбыкласс Osteichthyes — Костные рыбыподкласс Sarcopterygii — Лопастеперые рыбыотряд Coelacanthiformes — Целакантообразныеотряд Dipnoi — Двоякодышащие

класс Osteichthyes — Костные рыбы

подкласс Sarcopterygii — Лопастеперые рыбы
Неокостеневающая упругая хорда -основа осевого скелетател позвонков неточень подвижные и массивные или длинные опорные лопасти парных плавников снабжены особым внутренним скелетом из удлиненных костейопорный тазовый пояс брюшных плавников расположен вблизи клоакального выводного отверстиячешуя космоидного типаспиральный клапан в кишечнике, артериальный конус в сердце

oтряд Coelacanthiformes — Целакантообразные
Хвостовой плавник дифицеркальный из трех лопастейноздри внутренние, хоан нетлучей жаберной перепонки нетпередний спинной плавник расположен впереди от середины телакосмоидная чешуялегкое редуцировано
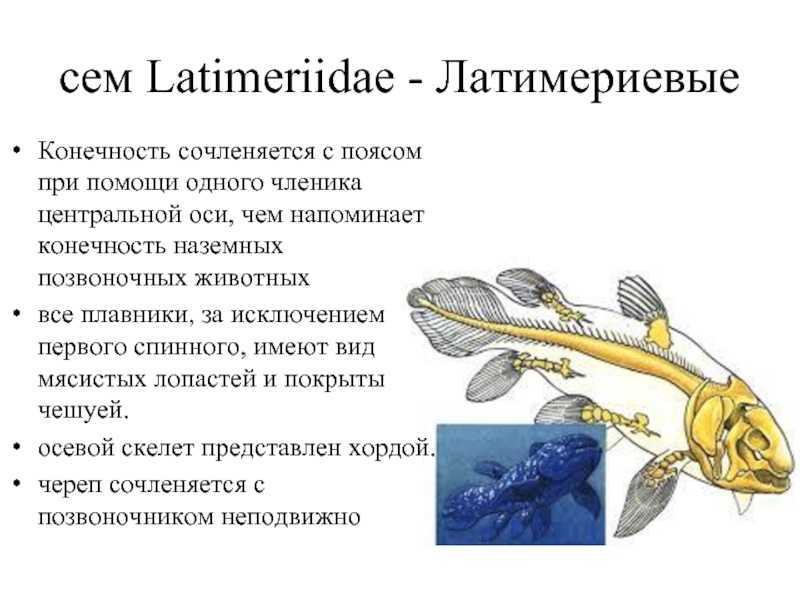
сем Latimeriidae — Латимериевые
Конечность сочленяется с поясом при помощи одного членика центральной оси, чем напоминает конечность наземных позвоночных животныхвсе плавники, за исключением первого спинного, имеют вид мясистых лопастей и покрыты чешуей. осевой скелет представлен хордой. череп сочленяется с позвоночником неподвижно

живородящий видокрашена в серо-синий цветдлинна 180 см, масса 95 кгморские, полуглубоководные хищникиактивны ночью
Latimeria chalumnae
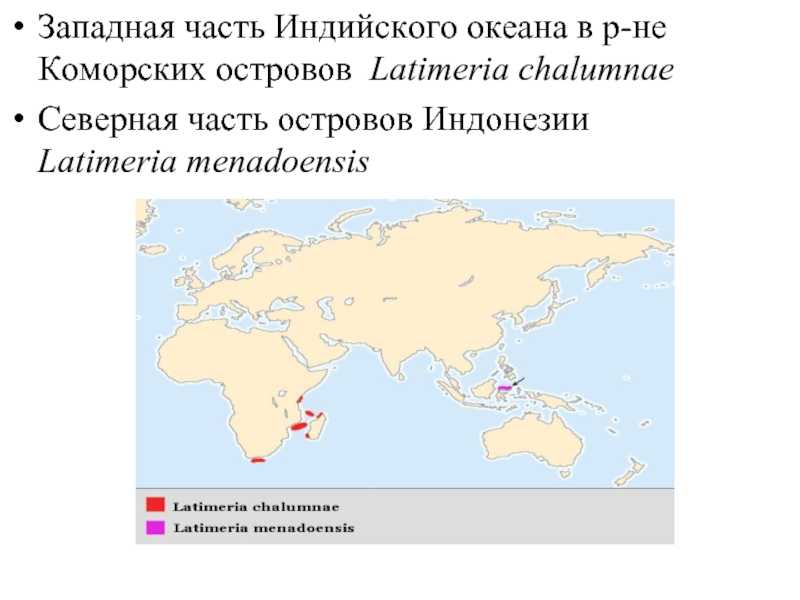
Западная часть Индийского океана в р-не Коморских островов Latimeria chalumnaeСеверная часть островов Индонезии Latimeria menadoensis

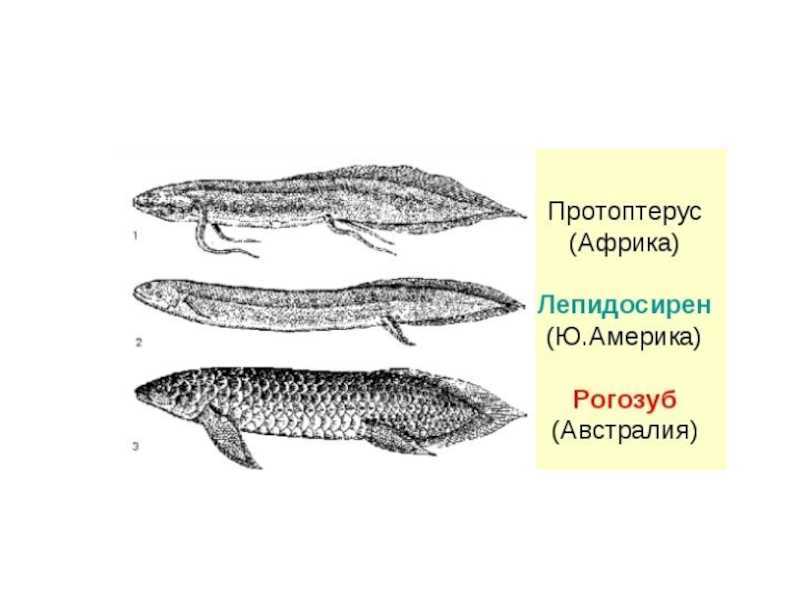
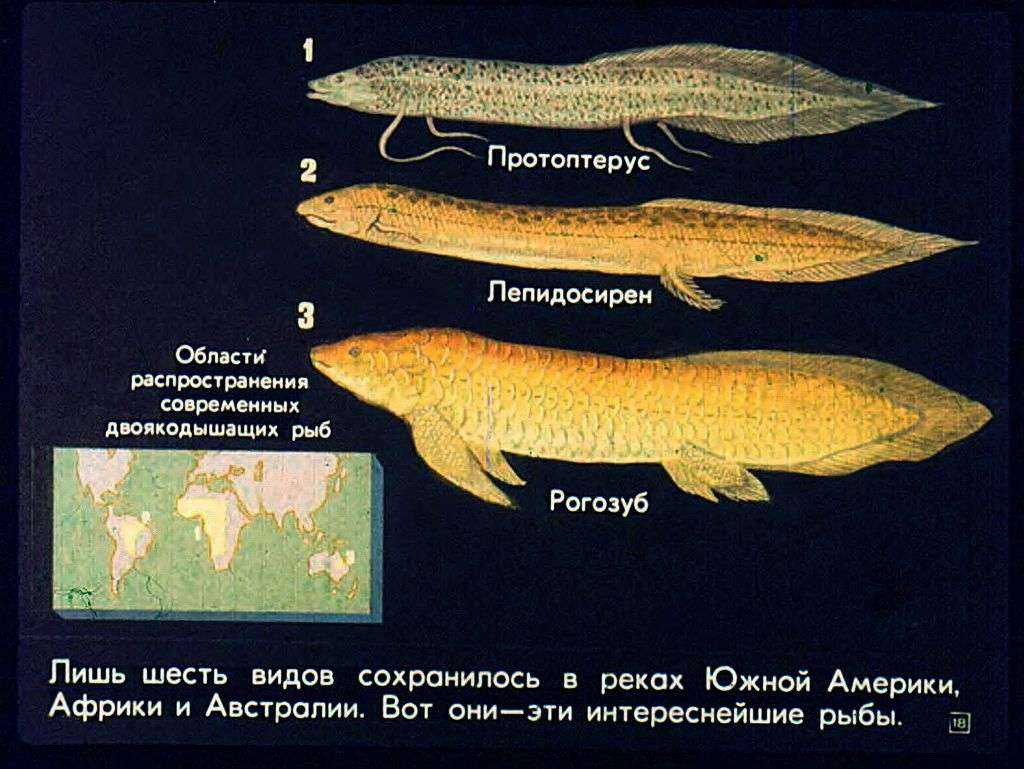
отряд Dipnoi- Двоякодышащие
Дышат жабрами растворенным в воде кислородом и атмосферным воздухомвнутренние ноздриплавательный пузырь имеет ячеистое строениев сердце — перегородки, есть легочный круг и задняя полая вена.чешуя циклоиднаяв скелете сохраняется много хрящаредуцированы межчелюстные, челюстные и зубные костив сердце сохраняется артериальный конус, в кишечнике – спиральный клапанесть клоакахвостовой плавник дифицеркальный

Центральная часть Южной Америки, Центральная Африка и Австралияпресноводные, обитающие в мелких заросших и пересыхающих водоемахактивны лишь в период дождей, а при пересыхании водоемов переходят на легочное дыханиеразмножаются откладыванием икрыпитаются беспозвоночными и рыбоймясо используется в пищу

сем Lepidosirenide – Чешуйчатниковые или Двулегочные
Угреобразное телоконечности в виде жгутов парные воздушные мешкиLepidosiren Paradoxa обитает в центральной части Южной Америки, а четыре других вида – в Центральной Африкемогут выносить полное пересыхание водоемов и впадать в спячку до 9 мес
Lepidosiren paradoxa
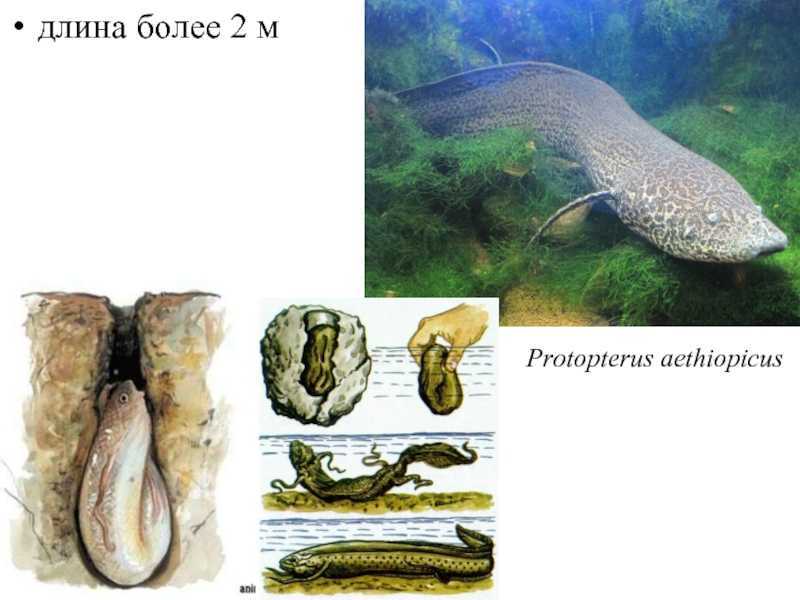
длина более 2 м
Protopterus aethiopicus

сем Ceratodidae – Рогозубовые, или Однолегочные
пресноводный видобитает в водоемах Австралиидлина тела 175 см, масса – 10 кгодно легкое и парные конечности в виде лопастейпитается различными беспозвоночнымиупотребляют в пищу
рогозуб, или неоцератод – Neoceratodus forsteri

СПАСИБО ЗА ВНИМАНИЕ!
легкие
Боковой вид легких рассеченной пятнистой двоякодышащей рыбы ( Protopterus dolloi )
Двоякодышащие рыбы имеют узкоспециализированную дыхательную систему . Они имеют отличительную особенность в том, что их легкие соединены с гортанью и глоткой без трахеи. В то время как другие виды рыб могут дышать воздухом с помощью модифицированных васкуляризированных газовых пузырей , эти пузыри обычно представляют собой простые мешочки, лишенные сложной внутренней структуры. Напротив, легкие двоякодышащих рыб подразделяются на множество более мелких воздушных мешочков, что максимально увеличивает площадь поверхности, доступную для газообмена.









Большинство существующих видов двоякодышащих рыб имеют два легких, за исключением австралийских двоякодышащих рыб, у которых только одно. Легкие двоякодышащих рыб гомологичны легким четвероногих . Как и у четвероногих и двуногих , легкие отходят от вентральной поверхности пищевода и кишечника.
Перфузия воды
Из существующих двоякодышащих рыб только австралийские двоякодышащие могут дышать через жабры, не нуждаясь в воздухе из легких. У других видов жабры слишком атрофированы, чтобы обеспечить адекватный газообмен . Когда двоякодышащая рыба получает кислород из жабр, ее кровеносная система устроена так же, как и у обычной рыбы. Спиральный клапан артериального конуса открыт, обходные артериолы третьей и четвертой жаберных дуг (фактически не имеющих жабр) закрыты, артериолы второй, пятой и шестой жаберных дуг открыты, артериальный протокотходящие от шестой артериолы открыты, а легочные артерии закрыты. Когда вода проходит через жабры, двоякодышащие рыбы используют буккальный насос. Поток через рот и жабры однонаправленный. Кровоток через вторичные ламеллы противоточен воде, поддерживая более постоянный градиент концентрации.
Перфузия воздуха
При дыхании воздухом спиральный клапан артериального конуса закрывается (минимизируя смешение оксигенированной и дезоксигенированной крови), открываются третья и четвертая жаберные дуги, закрываются вторая и пятая жаберные дуги (минимизируя возможную потерю кислорода, полученного в легких). через жабры), артериальный проток шестой артериолы закрывается, а легочные артерии открываются
Важно отметить, что при дыхании воздухом шестая жабра все еще используется при дыхании; деоксигенированная кровь теряет часть углекислого газа, проходя через жабры, прежде чем попасть в легкие. Это связано с тем, что углекислый газ лучше растворяется в воде
Поток воздуха через рот является приливным, а через легкие двунаправленным и наблюдается «равномерная бассейновая» диффузия кислорода.
Анатомия и морфология
Все двоякодышащие рыбы демонстрируют непрерывную хрящевую хорду и сильно развитый небный зубной ряд. Базальные (« примитивные ») группы двоякодышащих рыб могут сохранять маргинальные зубы и окостеневшую мозговую оболочку, но производные группы двоякодышащих рыб, включая все современные виды, демонстрируют значительное уменьшение маргинальных костей и хрящевой мозговой оболочки. Кости крыши черепа примитивных двоякодышащих рыб покрыты минерализованной тканью , называемой космином , но в постдевонскиеу двоякодышащих крыша черепа лежит под кожей, и космическое покрытие утрачено. У всех современных двоякодышащих рыб наблюдаются значительные редукции и слияния костей крыши черепа, а определенные кости крыши черепа не обнаруживают гомологии с костями крыши черепа лучеперых рыб или четвероногих . Во время сезона размножения у южноамериканской двоякодышащей рыбы развивается пара перистых придатков, которые на самом деле представляют собой сильно видоизмененные брюшные плавники. Считается, что эти плавники улучшают газообмен вокруг икры рыбы в ее гнезде.
В результате конвергентной эволюции у двоякодышащих рыб развились внутренние ноздри, похожие на хоаны четвероногих , и мозг, имеющий некоторое сходство с мозгом лиссамфибий (за исключением квинслендских двоякодышащих рыб, которые ответвились в своем направлении около 277 миллионов лет назад и мозг, напоминающий мозг латимерии ).
Зубной ряд двоякодышащих рыб отличается от зубного ряда любой другой группы позвоночныхОдонтоды » на небе и нижней челюсти развиваются в виде ряда рядов, образуя веерообразную окклюзионную поверхность. Затем эти зубы изнашиваются, образуя однородную поверхность дробления. У некоторых групп, в том числе у современных чешуеобразных , эти гребни были изменены, чтобы сформировать закрывающие лезвия.
Современные двоякодышащие рыбы имеют ряд личиночных особенностей, которые предполагают педоморфоз . Они также демонстрируют самый большой геном среди позвоночных.
Все современные двоякодышащие рыбы имеют удлиненное тело с мясистыми парными грудными и брюшными плавниками и одним непарным хвостовым плавником, заменяющим спинной, хвостовой и анальный плавники у большинства рыб.
Описание
У большинства лопастепёрых (включая и наиболее примитивных представителей класса) в парных плавниках имелись хорошо развитые мясистые лопасти, отличавшиеся расчленённым внутренним скелетом и наличием сложной мускулатуры (отсюда и происходят названия «лопастепёрые», «мясистолопастные»). У современных двоякодышащих конечности приобрели ланцетовидную или жгутовидную форму.
У древних лопастепёрых тело было покрыто толстыми ромбическими космоидными чешуйками, причём находившиеся между зубчиками косминового слоя и порами, открывавшимися наружу, полости вмещали, по-видимому, электрорецепторы. По мере эволюции происходила редукция косминового слоя, который полностью исчез у современных представителей лопастепёрых.
Древние лопастепёрые (в отличие от большинства лучепёрых) имели два спинных плавника (эта особенность характерна и для современной латимерии). Элпистостегалии утратили спинные плавники. Хвостовой плавник лопастепёрых изначально был гетероцеркальным, позднее стал дифицеркальным.
Опорно-двигательный аппарат: движение двоякодышащих производится с помощью боковых изгибов тела. С помощью плавников могут ползать по дну и переползать в другой водоём.
Дыхание: жабры и лёгкие. Механизм вентиляции: ротоглоточный и с помощью жаберных крышек.
У большинства лопастепёрых (кроме целакантообразных) парные наружные ноздри соединялись носовыми ходами с хоанами (внутренними ноздрями). Палеонтологи считают, что первоначальным назначением хоан было принудительное омывание сквозным потоком обонятельного эпителия.
Кровеносная система: поскольку имеются лёгкие, появляется второй (лёгочный, малый) круг кровообращения.
Пищеварение: разнообразные способы питания. Есть 3 отверстия: половое, анальное, выделительное.
Размножение: внутреннее оплодотворение, небольшое количество икринок могут развиваться в теле матери.
Нервная система: хорошо развит передний мозг, появляются хоаны (внутренние ноздри).
Двоякодышащие дышат жабрами или легкими — видоизмененный плавательный пузырь, соедененный с пищеводом. Есть основание парных плавников — мясистые, что позволяет опираться плавниками на грунт.
Дальнейшее чтение
- Альберг, ЧП; Смит, ММ; Йохансон, З. (2006). «Пластичность развития и несоответствие в зубных рядах ранних двоякодышащих (двоякодышащих) рыб». Эволюция и развитие . 8 (4): 331–349. doi10.1111/j.1525-142x.2006.00106.x . PMID . S2CID .
-
Палмер, Дуглас, изд. (1999). «Энциклопедия динозавров и доисторических существ Саймона и Шустера». Великобритания: Marshall Editions Developments Limited: 45.
Наглядное изображение доисторической жизни.
- Шульце, HP; Чорн, Дж. (1997). «Пермско-каменноугольный род Sagenodus и начало современных двоякодышащих рыб» . Вклад в зоологию . 61 (7): 9–70. doi10.1163/18759866-06701002 .
- Сепкоски, Джек (2002). «Сборник родов ископаемых морских животных» . Бюллетени американской палеонтологии . 364 : 560. Архивировано из оригинала 20 февраля 2009 г. Проверено 17 мая 2011 г.
Таксономия
Иллюстрация Цератода Генриха Хардера
Родство двоякодышащих рыб с остальными костистыми рыбами хорошо изучено:
- Двоякодышащие наиболее тесно связаны с Powichthys , а затем с Porolepiformes .
- Вместе эти таксоны образуют Dipnomorpha , сестринскую группу Tetrapodomorpha .
- Вместе они образуют рипидистию , сестринскую группу латимерий .
Недавний молекулярно-генетический анализ убедительно подтверждает сестринское родство двоякодышащих рыб и четвероногих ( Rhipidistia ), при этом латимерии разветвляются немного раньше.
Отношения между двоякодышащими рыбами разрешить значительно труднее. В то время как у девонских двоякодышащих рыб было достаточно костей в черепе, чтобы определять отношения, постдевонские двоякодышащие рыбы представлены полностью крышами черепа и зубами, так как остальная часть черепа хрящевая . Кроме того, многие из уже идентифицированных таксонов могут не быть монофилетическими .
Текущие филогенетические исследования подтверждают следующие взаимоотношения основных таксонов двоякодышащих рыб: класс Osteichthyes, подкласс Sarcopterygii, отряд Dipnoi.
| Двоякодышащие |
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
В 2017 году Энн Кемп , Лайонел Кальвин и Гийом Гино нашли другую классификацию более производных двоякодышащих рыб. В этом новом анализе все существующие двоякодышащие рыбы принадлежат к подотряду Ceratodontoidei , и, несмотря на их сходный внешний вид, нет отдельной клады, содержащей только Ceratodus , Neoceratodus и их ближайших союзников.










|
Rhinodipterus kimberleyensis † |
||||||||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||||||||
Новый анализ взаимоотношений недавно обнаруженного Persephonichthys chthonica и современных двоякодышащих рыб надежно относит оба таксона к двоякодышащим, а не к кладе, содержащей позднедевонские «фанероплевриды» и обычных позднепалеозойских двоякодышащих рыб, таких как Sagenodus . Монофилия постдевонских двоякодышащих рыб не поддерживается, и установлено, что каменноугольно-пермский таксон Sagenodus имеет случайное происхождение по отношению к происхождению современных двоякодышащих рыб, что свидетельствует о широкой конвергенции позднепалеозойских двоякодышащих рыб.
Лепидосирены
Единственный существующий на сегодня вид — американские чешуйчатники. Эти двоякодышащие рыбы похожи на другие виды, отличаются удлиненной формой тела и наличием пяти жаберных дуг и четырех щелей.
Чешуйчатники охотятся днем, передвигаясь по дну. Рацион состоит из моллюсков, мелкой рыбы и растений.

Половая зрелость наступает в 3-летнем возрасте. По истечению 3 недель после начала дождливого сезона начинается нерест. Самцы строят гнезда для выводка и защищают его. Икру откладывает самка через две недели. Желточный мешок рассасывается примерно через 30–40 дней. Молодняк сначала поедает водоросли и планктон.